

Autor : De Vito, Eduardo L.1-2
1Instituto de Investigaciones MĂŠdicas Alfredo Lanari, Facultad de Medicina, Universidad de Buenos Aires. 2 Centro del Parque, Cuidados Respiratorios, Buenos Aires, Argentina.
https://doi.org./10.56538/ramr.PGZO3337
Correspondencia : Eduardo L. De Vito E-mail: eldevito@gmail.com
RESUMEN
Este artículo analiza ciertos aspectos evolutivos
en el intercambio gaseoso, el desaÂrrollo pulmonar, la bomba respiratoria, el
estado ácido-base y el control de la ventilaÂción en
relación con un evento trascendente: el pasaje de la vida
acuática a la terresÂtre. Su estudio puede permitir comprender ciertos
aspectos con los que lidiamos en la práctica clínica: ÂżPor
qué las personas con debilidad muscular respiratoria extrema respiran
como ranas (respiración frog)?, ÂżPor
qué los recién nacidos con dificultad respiratoria tienen aleteo
nasal y quejido espiratorio?, Âżcómo es posible que los músÂculos
abdominales, típicamente espiratorios, asistan a la inspiración
en casos de la parálisis diafragmática?, Âżpor qué en la
insuficiencia respiratoria el patrón respiratorio tiene menos
variabilidad y se torna más rígido? y, por último, Âżes
posible imaginar un pH neutro que no tenga el valor de 7,0, para qué
sirve este conocimiento y como se deben interpretar los gases en hipotermia?
La transición del agua a la tierra es una de las
más importantes e inspiradoras de las grandes transiciones en la
evolución de los vertebrados. Ante la sorprendente diversiÂdad de
organismos vivos, es tentador imaginar una cantidad enorme de adaptaciones
evolutivas para resolver los diferentes desafíos que cada especie tiene
para la vida en la tierra. Hay desarrollos tempranos que comparten algunos
factores cruciales y algunas de las redes genéticas regulatorias
cercanas y lejanas están conservadas. Somos testigos de hallazgos
clínicos que son el testimonio de especies que han vivido en
épocas remotas y nos han legado su historia evolutiva.
Palabras clave: Equilibrio ácido-base; Hipotermia; Imidazol;
Evolución biológica; Parálisis respiratoria; Centro
respiratorio
ABSTRACT
This article analyzes certain
evolutionary aspects of gas exchange, lung development, the respiratory pump,
the acid-base status and control of ventilation in relation to a significant
event: the passing from aquatic to terrestrial life. By studying this, we can
understand certain aspects that are present in the clinical practice: Why do
people with extreme respiratory muscle weakness breathe as frogs? (frog
breathing); why do newborns with breathing difficulties have nasal flaring and
expiratory grunting?; how is it possible that abdominal muscles, which are
typically expiratory, assist with inspiraÂtion in cases of diaphragmatic
paralysis?; why does the breathing pattern of respiratory failure has less
variability and becomes more rigid? and, finally, is it possible to imagine a
neutral pH that doesnât have the 7.0 value?; whatâs the use of this knowledge,
and how should gases in hypothermia be interpreted?
Water-to-land transition is one
of the most important and inspiring major transitions of vertebrate evolution.
Given the amazing diversity of living organisms, it is tempting to imagine an
enormous amount of evolutionary adaptation processes to solve the different
challenges of living on earth faced by each species. There are certain early
development processes that share some crucial factors, and some of the close
and distant gene regulatory networks are conserved. We are witnesses of clinical
findings that serve as testimony of the species that lived in remote times and
left us their evoÂlutionary history.
Key words: Acid-base equilibrium; Hypothermia; Imidazole; Biological evolution;
Respiratory paralysis; Respiratory center
Recibido: 20/06/2022
Aceptado: 09/08/2022
Plus ça change,
plus câest la même chose.
Alphonse Karr (1808-1890)
El objetivo de este artículo es analizar ciertos
aspectos respiratorios evolutivos, particularmente en el intercambio gaseoso,
el desarrollo pulmonar, la bomba respiratoria, el estado ácido-base y el
control de la ventilación en relación con un evento trascendente:
el pasaje de la vida acuática a la vida terrestre.
Su estudio puede ayudar a comprender ciertos aspectos con
los que lidiamos frecuentemente en la práctica clínica: ÂżPor
qué las personas con debiÂlidad muscular respiratoria extrema respiran
como ranas (respiración frog)?, Âżpor
qué los recién naciÂdos con dificultad respiratoria tienen aleteo
nasal y quejido espiratorio?, Âżcómo es posible que los músculos
abdominales, típicamente espiratorios, puedan asistir a la
inspiración en casos de parálisis diafragmática?, Âżpor qué
en la insuficiencia respiÂratoria el patrón respiratorio (además
de rápido y superficial) tiene menos variabilidad y se torna más
rígido? y, por último, Âżes posible imaginar un pH neutro que no
tenga el valor de 7,0, para qué sirve este conocimiento y como se deben
interpretar los gases en hipotermia?
La transición del agua a la tierra es una de las
más importantes e inspiradoras de las grandes transiciones en la
evolución de los vertebrados. El primer pez apareció hace unos
438 millones de años y la transición del pez tetrápodo
hacia la tierra fue hace unos 375 millones de años; ellos fueron los
protagonistas de ese evento único: emergieron del agua y respiraron
aire. Eran exoÂtérmicos e incapaces de sostener niveles elevados de
actividad física y evolucionaron a dos grandes clases de vertebrados con
altos niveles de consumo máximo de oxígeno: los mamíferos
y las aves.1La habilidad
terrestre parece coincidir con el origen de las extremidades; hubo una
coexistencia de rasgos acuáticos, como las branquias, la aleta de la
cola y las extremidades.2
CAMBIOS EN LA COMPOSICIÓN DE GASES EN LA BIOSFERA
Las fluctuaciones de los niveles de O2 y CO2
en la biosfera han determinado las formas y los medios por los
cuales se incorporaba el O2 y
se eliminaba el CO2.
Durante la era Paleozoica tardía (unos 300 millones de años),
durante un período de unos 120 millones de años, el nivel de O2 aumentó
a un máximo del 35 % y, luego, cayó precipitadamente a un
mínimo del 15 % en el Triásico. Estos cambios se duplicaron en el
agua y tuvieron como resultado grandes eventos, como extinciones masivas.
Los niveles más altos de CO2 ocurrieron en el Ordovícico y el
Silúrico, mientras que, en el Carbonífero, ese nivel había
descendido al actual (0,036 %), aunque al final del Pérmico había
auÂmentado por un factor de tres. La estructura y función de los
intercambiadores de gases iniciales se produjeron en gran medida por
selección natural en condiciones ambientales que eran totalmente
diferentes a las actuales.3
EL AMBIENTE ACUOSO Y TERRESTRE
La composición gasométrica del aire es bien
conociÂda por nosotros. El oxígeno del agua de mar deriva en gran parte
del aire, por lo que está formado por los mismos gases de la
atmósfera. Como el oxígeno es más soluble en el agua que
el nitrógeno, se encuentra en mayor proporción que en el aire.
Pero desde el punto de vista del oxígeno disuelto (molecular), mientras
que en el aire hay 210 cmÂł de O2/L,
en el agua de mar hay solo 9 cmÂł/L. Así, en términos generales,
el oxígeno disuelto es mucho menos abundante en el agua que en el aire.
La presencia de algas macroscópicas y microsÂcópicas
contribuye en relación directa con la iluÂminación a la
oxigenación del agua de mar. Solo el 1 % de la luz que incide sobre la
superficie del mar llega a los 200 m de profundidad (zona fótica).4 Así
la disponibilidad de O2 disminuye
notablemente conforme aumenta la profundidad del agua.
INTERCAMBIO GASEOSO
La modificación más dramática en el
intercambio gaseoso se produjo como consecuencia de la modiÂficación en
la estructura de los tegumentos. En el agua, el intercambio gaseoso se
producía por dos vías: los tegumentos y las estructuras
respiratorias incipientes. Con el salto a la vida terrestre y la apaÂrición
de las escamas (reptiles) los tegumentos iban a proteger de la
desecación, pero se harían menos permeables al intercambio de los
gases.
En las aves y mamíferos, las plumas y los pelos
vedaron definitivamente el intercambio gaseoso cutáneo, función
que desempeñaría exclusivamenÂte el pulmón mediante el
ingreso de aire por la boca. Comenzó a conformarse el aire alveolar, una
estación intermedia entre el aire atmosférico y la sangre, con
una composición gasométrica, tempeÂratura y humedad
remarcablemente estables. De manera que el pulmón de los
mamíferos evolucionó para enfrentar un conjunto único de
desafíos:
â Asegurar un suministro eficiente de aire insÂpirado a
todas las unidades pulmonares cuya superficie de intercambio gaseoso
alcanzaría unos 70-150 m2en el hombre,
dentro de un espacio torácico confinado,
â esa gran superficie de intercambio de gases deÂbería
estar asociada con un espesor de barrera mínimo y,
â debería generarse una red microvascular
para acomodar todo el gasto cardíaco del ventrículo derecho y
resistir las tensiones mecánicas cícliÂcas que aumentan varias
veces desde el reposo hasta el ejercicio.
Los órganos del aparato respiratorio que tienen
diversos animales acuáticos, entre ellos los peces, son las branquias (o
agallas) y el opérculo. La dinámica de apertura y cierre de las
branquias está controlada por los nervios craneanos deriÂvados de los
arcos branquiales (trigémino, facial y glosofaríngeo). En las
ranas, la inervación del área branquial se desarrolla a partir de
los nervios craneales facial, glosofaríngeo y vago.5
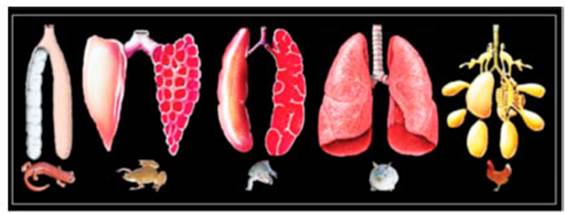
La Figura 1 muestra el aspecto morfológico de los pulmones
de los vertebrados según JN Marina.3
Los mamíferos y las aves son las dos grandes
clases de vertebrados con altos niveles de consumo máximo de
oxígeno.1Una
característica notable de estos dos grupos es que, aunque la
fisiología de los sistemas cardiovascular, renal, gastrointestinal,
endocrino y nervioso muestra muchas similitudes, los pulmones son radicalmente
diferentes.6
Nuestra perspectiva de mamíferos autoconsÂcientes
podría hacernos pensar que hemos sido más exitosos que las aves.
Desde ciertos aspectos parece ser lo opuesto. West y Watson propusieron que el
pulmón de las aves es superior al del mamífero, y que la
evolución fue por el camino equivocado para este último:1
⢠Una diferencia importante es que la ventilaÂción
de la zona de intercambio de gases (zona respiratoria de West) tiene un
patrón de flujo continuo en las aves, pero es alternante en el
mamífero.
⢠Las aves movilizan los gases mediante la convecÂción,
mientras que los mamíferos necesitan, adeÂmás, de difusión
en las vías aéreas terminales.7
⢠Las aves tienen un parénquima más
uniforme con espacios terminales pequeños muy entraÂmados con los
capilares, un espesor mínimo de la membrana y en definitiva un
intercambio de gases más eficiente.
⢠Las aves han separado las funciones de ventiÂlación
y de intercambio de gases, parecen ser menos vulnerables a broncoaspiración
y su consumo de oxígeno en relación con su peso corporal es mayor
que el de los mamíferos.
Por todo esto, desde el punto de vista de la estrucÂtura
y la función, el pulmón de las aves es superior.1
El hombre no fue la meta de la evolución (que no tiene
meta) y menos aún el pulmón de los mamíÂferos. La
evolución procede en forma gradual, no necesariamente hacia estructuras
más complejas. 8
En virtud del gran desarrollo de su cerebro, el hombre es
una criatura agudamente autoconscienÂte, capaz, en un grado inmensamente mayor
que cualquier otro animal, de aprovechar la experiencia individual y social.
Pero mientras que un alpinista lucha para llegar a la cima del Everest, lo
están esperando unos gansos volando sobre su cabeza.
LA BOMBA RESPIRATORIA
La evolución a la vida terrestre confinó el
intercamÂbio gaseoso a los pulmones que evolucionaron hacia una gran superficie
de intercambio expuesta a un aire alveolar muy controlado que hubo que mover
(ventilar) para tomar aire desde la atmósfera. La bomba respiratoria en
sus diferentes versiones se encargó de esto último.9
La Figura 2 muestra una representación
gráfica (dendrograma) de varios grupos de
vertebrados en relación con la estrategia utilizada respecto de la bomba
respiratoria.9 Es posible
observar el cambio de una bomba bucal impulsada por músculos branÂquioméricos (del aparato faríngeo) e hipobranquiales (laringe, lengua, mandíbula)
inervados por nervios craneales a una bomba de aspiración torácico-abdominal
impulsada por músculos axiales inervaÂdos por nervios espinales con
neuronas premotoras situadas en la columna
respiratoria ventral.
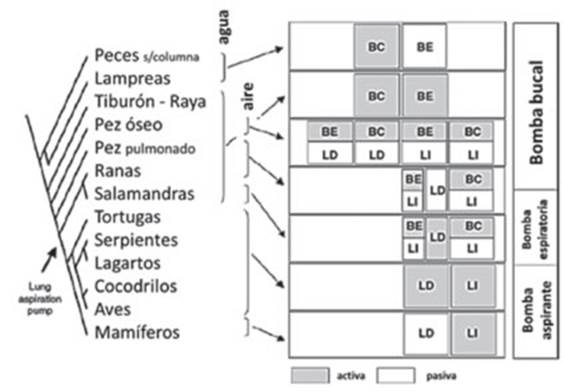
Los pasos iniciales en la evolución de la respiraÂción
en el aire fueron una modificación del comporÂtamiento en la superficie
y cambios en las válvulas de la boca/espiráculo/narinas y el
opérculo y la glotis (o sus equivalentes), es decir, cambios en la
activación de los músculos que dilatan o contraen varias
aberturas. Esto permitió tanto la respiración acuática
como aérea. Los cambios en los músculos de la bomba respiratoria
evolucionaron más tarÂde.10De manera que
la evolución de los mecanismos respiratorios en los vertebrados progresa
desde la ventilación acuática promovida principalmente por una
bomba de fuerza bucal hasta la ventilación aérea impulsada
principalmente por una bomba de aspiración o succión. Solo
los mamíferos tienen diafragma muscular, su origen es axial y
está inervado por neuronas motoras espinales (nervio frénico) y
no por los pares craneanos.
Entre ambos extremos de la bomba bucal y bomba aspirante
tuvo lugar la espiración activa (Figura 2, bomba espiratoria).9Este
mecanismo intermedio entre la bomba bucal de peces y anfibios y la bomba de
aspiración de reptiles, aves y maÂmíferos era poco conocido. Se
ha demostrado que muchos anfibios usan los músculos axiales para la
espiración activa junto con la bomba bucal para la inspiración
activa. Esto sugiere que la respiración por aspiración
evolucionó en dos pasos:
⢠del bombeo bucal exclusivo, al bombeo bucal para la
inspiración y los músculos axiales para la espiración y,
luego, tuvo lugar
⢠la respiración por aspiración exclusiva,
usando los músculos axiales tanto para la espiración como para la
inspiración.
En los mamíferos, vemos un cambio en las
contribuciones relativas de la distensibilidad de la
pared torácica y la resistencia del flujo de aire al trabajo pulmonar
(la primera domina en aves y reptiles, la última adquiere mayor
importancia en los mamíferos). Además, se observa la
evolución a un diafragma muscular y una reducción en la
necesidad de deflación pulmonar activa a meÂdida que el sistema vuelve
al estado de equilibrio después de la inhalación (retroceso
elástico). Se va formando así la capacidad residual funcional
(CRF), un volumen pulmonar al fin de la espiración tranquila y que representa
el balance entre las fuerzas que expanden el pulmón y las que tienden a
colapsarlo (reposo respiratorio).
En peces, anfibios y la mayoría de los reptiles,
existe una cierta división entre la cavidad torácica y la
abdominal. Esta partición es incompleta y muy poco eficiente como bomba
respiratoria. En los reptiles avanzados y en todos los mamíferos, se
consigue una separación completa de las caviÂdades torácica y
abdominal; esta separación se musculariza en
el caso de los mamíferos. Estamos en presencia del diafragma como músculo
y de la respiración por aspiración (presión negativa).10
EVOLUCIÓN DE LA REGULACIÓN ÁCIDO BASE
En los seres humanos, la PaCO2 es controlada de manera estricta. A lo largo
del día y de la noche y aún con la participación de otras
funciones no respiratorias, su variación es de unos pocos mmHg. Además, a diferencia de la PaO2 que declina con la edad, la PaCO2 permanece
constante durante toda la vida. Por esto, cualquier desviación sostenida
de la PaCO2 debe
ser vista como una alteración significativa de la homeostasis.11
ÂżCuál es la importancia del control tan estricto
de la PaCO2 y
cómo se logra? El estudio de la evoÂlución de los vertebrados
desde la vida acuática hacia la terrestre y de la capacidad de regular
la temperatura corporal, permiten entender por qué la PaCO2 debe ser
mantenida dentro de límites tan estrechos.
En la vida acuática bajo una
dinámica poiquiÂloterma, la temperatura corporal no es constante, sino
que varía según la temperatura ambiente. La PCO2 sufre grandes variaciones y, si bien su elimiÂnación
es muy sencilla (tegumentos permeables al CO2), el problema principal es la
oxigenación (la PO2 del
agua es más baja que la atmosférica) y, por diversas razones que
veremos, resulta imposible mantener un valor constante de pH.
En la vida terrestre, en cambio, los quimioÂrreceptores
periféricos (QRP) dejan de funcionar debido a la alta PO2 ambiental (son sensibles a la PaO2 por debajo
de 60 mmHg), la temperatura corÂporal se puede
mantener constante (homeotermia), pero ahora la
única vía de eliminación del CO2 es el aire espirado (debido al desarrollo de
tegumentos que evitan la desecación).
Con una capacidad de regulación térmica
bien desarrollada y con la precisa regulación de la PaCO2, el pH resultante, remarcablemente estable,
permite a los mamíferos mantener la ionización de las enzimas y
productos del metabolismo intermeÂdio y no escapar de la célula (las
enzimas escapaÂrían de la célula si perdieran su
ionización). Esta estrategia se denomina pH-stat,
es decir, resulta muy importante para los homeotermos mantener el pH constante
(límites muy estrechos). Según esto, los intermediarios
metabólicos y enzimas están completamente ionizados en la
región cerÂcana al pH neutro (pH neutro 7,0 a 25 °C) y tienen poca
tendencia a escapar de la célula atravesando las membranas.
En otros términos, si el pH del medio se apartara
de la ventana de ionización de los intermediarios metabólicos,
estos dejarían de tener carga y esÂcaparían de la célula.
De ahí la importancia de la constancia del pH (pH-stat).
Esto fue eleganteÂmente expresado como âla importancia de estar ionizadosâ.12
Para entender por qué esta estrategia no es efecÂtiva
en los poiquilotermos, conviene puntualizar un hecho pocas veces tenido en
cuenta dado que la temperatura corporal de 37 °C no nos plantea la
relación entre la temperatura y el pH (excepto en puntuales ocasiones
como en la hipotermia accidental o terapéutica): La temperatura
cambia el valor de neutralidad del pH debido a cambios en la constante de
equilibrio del agua o Kw (proÂducto iónico del
agua). Así, el agua pura tiene un pH neutro (valor 7,0) solo a 25 °C,
mientras que a 10 °C y a 35 °C el valor de pH neutro es de 7,27 y 6,98,
respectivamente*.
En el agua, a bajas temperaturas, el pH del meÂdio
interno de los poiquilotermos (peces, anfibios, reptiles) tiende a aumentar y
como consecuencia, se aleja de la ventana de ionización de las
proteínas y enzimas que, en tanto buffers pueden perder ioÂnización,
pero afortunadamente la histidina con su grupo α-imidazol
conserva su ionización constante; así, esa ionización
mantiene a las enzimas dentro de la célula y activas, no obstante, las
variaciones en la temperatura. Esta es la denominada estraÂtegia α-stat.12-15
La histidina es un aminoácido muy particular.
Tiene tres grupos capaces de adquirir carga: amino (pK
9,17), carboxilo (pK 1,82) e imidazol
(pK 6,0) y su carga neta (o grado de
disociación) permanece constante en todo el rango de temperatura y es la
base de la teoría α-stat de Reeves.15
En realidad los poiquilotermos
acuáticos estuÂvieron mucho antes que los homeotermos terrestres y
fuimos nosotros los que tuvimos que hallar una estrategia homeostática
cuando los tegumentos perdieron la permeabilidad al CO2. La constancia de
la PCO2 es
una conquista de la vida terrestre y quizá no hubiese tenido lugar sin
la dramática complejidad que desarrollaron las estructuras controladoras
de la respiración.
Pero la constancia del pH en humanos es el resultado
también de la interacción de múltiples sistemas buffer en
los que se encuentran los sisteÂmas proteicos y la regulación precisa
del sistema bicarbonato/ácido carbónico mediante el control
ventilatorio y renal. Es evidente que todo esto ha sido posible debido a la
evolución de los centros respiratorios.
EVOLUCIÓN DEL CONTROL DE LA VENTILACIÓN
En el ambiente acuático, los QRP de los poiquiloÂtermos
se encargan de regular minuto a minuto la ventilación así como el
nivel de inmersión. Sus centros respiratorios consisten en grupos de
céluÂlas relativamente sencillas, capaces de generar un patrón
respiratorio muy simple; por ejemplo, los anfibios utilizan solo dos grupos de motoneuronas que median la ventilación. Lejos
aún de adquirir el nervio frénico, los primeros nervios
involucrados en el acto de la respiración fueron el facial y el
glosofaríngeo.
Con el salto a la vida terrestre, las estructuras
generadoras del ritmo respiratorio son ahora redes neurales oscilatorias de
seis grupos de motoneuronas interconectadas y se
desarrollan quimiorreceptores sensibles al CO2. Los nuevos circuitos neuronales son estables
pero responÂdedores a cambios en los niveles de O2, CO2,
pH, ejercicio, sueño, etc. Además, debe haber una
coordinación con la fonación, deglución, reflejos de la
vía aérea, tos, estornudo, locomoción. A esto se suma la
adaptación a largo plazo por alÂteraciones de la caja torácica,
del pulmón y de los músculos respiratorios con la edad, la
ganancia o la pérdida de peso, el embarazo y las enfermeÂdades.
Finalmente, las nuevas estructuras supraÂpontinas
controlan los músculos respiratorios en forma voluntaria y en
relación una âcuriosidad evolutivaâ: las emociones.
Si es que hay algo que ya forma parte de la historia de
la medicina es el modelo de centros respiratorios en mamíferos con los
núcleos pneuÂmotáxico, apnéustico y grupo respiratorio dorsal y grupo
respiratorio ventral. Este modelo surgió de cortes transversales del
tronco de gatos anesÂtesiados, descerebrados a la altura intercolicular,
desaferentizados, con vago o sin este y de la obÂservación
de los cambios de patrón respiratorio y, luego, de un total de seis transecciones a diferentes niveles del tronco.16, 17
En la actualidad, no es posible abordar el tema de los centros
respiratorios si no se menciona al complejo preBötzinger
como un área crítica para la generación del ritmo
respiratorio, y de núcleo retrotrapezoidal y
del grupo respiratorio parafacial para la
generación de la espiración activa y para la relación con
funciones no respiratorias.17-20La Figura 3
muestra la organización esquemática del sistema respiratorio.
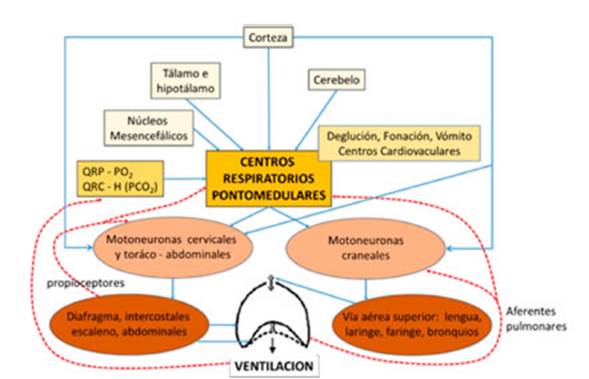
De manera que la evolución del control respiraÂtorio
desde peces a mamíferos se caracteriza por el aumento de la complejidad
y está relacionada con la homeostasis del O2, CO2,
pH y la temperatura.
â Los QRP sensibles a la hipoxia (PaO2 ≤ 60 mmHg y
muy activos bajo el agua) dejaron de funcionar en la tierra.
â Cuando se priorizó la constancia de la PaCO2 y del pH
(estrategia pH-stat) se desarrollaron
áreas muy sensibles al CO2.
â Las simples células marcapasos dieron lugar al
complejo preBötzinger, así como el grupo
respiÂratorio parafacial y el núcleo
retrotrapezoidal.23,
24
â Se desarrollaron conexiones con funciones no
respiratorias para coordinar respiración con fonación,
deglución y vómito.25
â Tenemos una suerte de homúnculo (área
motora primaria de la corteza, lóbulo frontal posterior,
circunvolución precentral) que es capaz de
comandar voluntariamente la respiración por sobre el control
autonómico.26
â La telencefalización
alcanzó su máxima expreÂsión en humanos. La corteza y las
estructuras subcorticales-sistema límbico (la amígdala, la
circunvolución cingulada, el hipocampo, etc.)
median las emociones e influencian la respiraÂción.
â La red respiratoria pontomedular,
que produce el comando central rítmico para la respiraÂción,
conforma estructuras estables, coordinaÂdas y adaptables.
CONTROL DE LA VENTILACIÓN Y ESTADO ÁCIDO-BASE. LECCIONES
CLÍNICO-EVOLUTIVAS
Solo en los reptiles avanzados y en todos los maÂmíferos,
se consigue una separación completa de las cavidades torácica y
abdominal; esta separaÂción se musculariza en
el caso de los mamíferos: el músculo diafragma. Se evoluciona
así hacia la respiración por aspiración, patrimonio
de maÂmíferos terrestres y acuáticos, pero nuestro acervo
genético parece recordar otros pasos evolutivos.
â Algunas personas con enfermedades neuroÂmusculares y
marcada debilidad muscular respiratoria utilizan la respiración frog, que permite el ingreso de aire a
presión positiva. La respiración frog,
a presión positiva (bomba bucal), constituyó una de las
primeras modaÂlidades ventilatorias de los vertebrados.
â El aleteo nasal y el quejido espiratorio son
signos de dificultad respiratoria en el recién nacido,27
bebés y niños pequeños.28
Indican aumento del esfuerzo para respirar. Este meÂcanismo es
inusual en adultos.28 Diversos
pares craneanos coordinados mueven esas estructuras y recuerdan la
respiración con válvulas de la boca/espiráculo/narinas
y el opérculo y la glotis (o sus equivalentes) de los primeros
vertebrados.
â En presencia de parálisis diafragmática
bilateral los músculos abdominales tienen acción inspiratoria.
Su acción reduce la CRF, así el inicio de la próxima inspiración
el aire ingresa en forma pasiva por la vuelta al equilibrio del tórax. Los
músculos abdominales (bomba espiratoria) tienen función
inspiratoria en ciertos vertebrados.
â La variabilidad de patrón respiratorio es
menor en presencia de insuficiencia respiratoria aguda. La utilización
de ventilación no invasiva reestablece la
variabilidad y se acerca a la norÂmalidad con niveles más altos de
soporte.29El
patrón respiratorio rígido, con poca variabilidad recuerda a los
poiquilotermos. La evolución ganó en complejidad y variabilidad,
pero, ante un aumento de la carga, el patrón respiratorio se hace
más rígido.
La utilización de hipotermia corporal general para
la cirugía cardíaca ha pasado a ser proceÂdimiento rutinario.
Esto obligó a repensar el concepto de pH neutro y atender a la
experiencia de millones de años de nuestros ancestros los
poiquilotermos.
â La definición de neutralidad (que se
remonÂta a Arrhenius, 1889): no es âun pH de 7,0â, sino la
presencia de cantidades iguales de iones H+ y OHâ.
Debido a que la temperatura tiene efecÂtos iguales en la
concentración de cada uno de ellos, la neutralidad se conserva sin
importar la temperatura.30
â Independientemente de la temperatura del paÂciente, los
gases arteriales se analizan siempre a 37 °C (es la temperatura con la que
miden los electrodos de PO2,
PCO2 y
pH). Los gases del paciente hipotérmico, también son analizados a
37 °C y si los valores de PO2,
PCO2 y pH
se encuentran dentro del rango normal, el estado ácido-base del paciente
será el adecuado para su temperatura.31, 32
â La sangre de un mamífero in vitro,
enfriada anaeróbicamente sigue el patrón de estado áciÂdo-base
de un poiquilotermo. Corregir el pH por la temperatura corporal en
hipotermia no tiene sentido fisiológico, ya que el pH de neutralidad
también cambia con la temperatura.
Ante la sorprendente diversidad de organismos vivos, es
tentador imaginar una cantidad enorme de adaptaciones evolutivas para resolver
los diÂferentes desafíos que cada especie tiene para la vida en la
tierra. Hay desarrollos tempranos que comparten algunos factores cruciales y
algunas de las redes genéticas regulatorias cercanas como lejanas
están conservadas.
En la Francia previa a la guerra franco-prusiana, en tono
de sátira política ante los cambios de gobierno se decía
âTomamos lo misÂmo y comenzamos de nuevoâ. Alphonse Karr
(1808-1890)33 sumó
su célebre âplus ça change, plus câest la même choseâ: cuanto
más cambian las cosas, más permanecen igual. En
biología comparada, parece haber principios inmutables aun con
diferencias superficiales o morfológicas evidentes. Es conmovedor
meditar que somos los únicos testigos de hallazgos clínicos que
son el testimonio de especies que han vivido en épocas remotas y nos han
legado su historia evolutiva; nuestra historia evolutiva.
Conflicto de intereses
El autor no tiene conflicto de intereses.
Notas
* Si el pH aumenta a medida que baja la temperatura, esto
no significa que el agua se vuelva más alcalina a temperaturas
más bajas. Una solución es alcalina si hay un exceso de iones
hidroxilo sobre iones hidrógeno (es decir, pOH
> pH). Siempre que haya la misma concentración de iones de
hidrógeno e iones de hidróxido, el agua sigue siendo neutra (pH =
pOH), incluso si su pH cambia. El problema es que
todos estamos familiarizados con que 7,0 sea el pH del agua pura (no ionizada)
y cualquier otra cosa se siente realmente extraña. Para calcular el
valor de pH neutro es necesario conocer el Kw âque
aumenta con la temperaturaâ y, si cambia, entonces el valor de pH neutro
también cambia. A 25 °C el valor de Kw (mol2 dmâ6) es de 1,00
x 10â14, el
pH es de 7,00 y el pOH es de 7,00. Así, 7,00 +
7,00 = 14. A 10 °C tendremos Kw 0,681 x 10â14, pH 7,08 y
pOH 7,08. Así, 7,08 + 7,08 = 14,16. Entonces
el pH 7,00 y 7,08 a 25 °C y a 10 °C, respectivamente, es neutro porque tiene H+ = OHâ.34
BIBLIOGRAFÍA
1. West JB, Watson RR, Fu Z. The
human lung: did evoluÂtion get it wrong? Eur Respir J 2007;29:11-17.
https://doi.org/10.1183/09031936.00133306
2. Dickson BV, Clack JA, Smithson
TR, et al. Functional adapÂtive landscapes predict terrestrial capacity at the
origin of limbs. Nature 2021;589:242-5.
https://doi.org/10.1038/s41586-020-2974-5.
3. Maina JN. Comparative
respiratory physiology: the funÂdamental mechanisms and the functional designs
of the gas exchangers. Open Access Animal Physiology 2014;53.
https://doi.org/10.2147/OAAP.S53213
4. Knoll AH, Bergmann KD, Strauss
JV. Life: the first two bilÂlion years. Philos Trans R Soc Lond B Biol Sci
2016;5:371. https://doi:10.1098/rstb.2015.0493.
5. Alonso DG. Desarrollo y diferenciación de las
branquias externas e internas en embriones y larvas de Bufo arenaÂrum:
análisis descriptivo y experimental. Tesis de Doctor. Facultad de
Ciencias Exactas y Naturales. Universidad de Buenos Aires. 2003.
http://digital.bl.fcen.uba.ar/Download/Tesis/Tesis_3581_Alonso.pdf
6. Hsia CCW, Hyde DM, Weibel ER. Lung Structure and
the Intrinsic Challenges of Gas Exchange. Compr Physiol 2016;6:827-95.
https://doi.org/10.1002/cphy.c150028.
7. West JB, Watson RR, Fu Z. The
human lung: did evoluÂtion get it wrong? Eur Respir J 2007;29:11-7.
https://doi.org/10.1183/09031936.00133306.
8. Smith HW. From Fish to
Philosopher. The American Biology Teacher 1962;24::445.
https://doi.org/10.2307/4440034
9. Milsom WK. Evolutionary Trends
in Respiratory MechaÂnisms. In: Poulin, M.J., Wilson, R.J.A. (eds) Integration
in Respiratory Control. Advances in Experimental Medicine and Biology 2008;605.
Springer, New York, NY. https://doi.org/10.1007/978-0-387-73693-8_51.
10. Fogarty MJ, Sieck GC.
Evolution and Functional DifferentiÂation of the Diaphragm Muscle of Mammals. Compr Physiol
2019;9:715-66. https://doi.org/10.1002/cphy.c180012.
11. Arce SC, De Vito EL. More Breathing, Less Fitness: LesÂsons from Exercise Physiology in
Chronic Obstructive Pulmonary DiseaseâHeart Failure Overlap. Am J Respir Crit
Care Med 2017;196:1233-4. https://doi.org/10.1164/rccm.201707-1430ED
12. Rahn H, Reeves RB, Howell BJ.
Hydrogen ion regulaÂtion, temperature, and evolution. Am Rev Respir Dis
1975;112:165-72.
13. Wilson RJA, Vasilakos K,
Remmers JE. Phylogeny of verteÂbrate respiratory rhythm generators: the
Oscillator HomolÂogy Hypothesis. Respir Physiol Neurobiol 2006;154:47-60. https://doi.org/10.1016/j.resp.2006.04.007
14. Rahn H. Body temperature and
acid-base regulation. Pneumonologie 1974;151:87-94.
https://doi.org/10.1007/BF02097155
15. Reeves RB. An imidazole
alphastat hypothesis for vertebrate acid-base regulation: Tissue carbon dioxide
content and body temperature in bullfrogs. Respir Physiol 1972;14:219- 36.
https://doi.org/10.1016/0034-5687(72)90030-8
16. Lumsden T. Observations on
the respiratory centres in the cat. J Physiol 1923;57:153-60. https://doi.org/10.1113/jphysiol.1923.sp002052
17. Stella G. On the mechanism of
production, and the physiÂological significance of âapneusisâ. J Physiol 1938;93:10-23.
https://doi.org/10.1113/jphysiol.1938.sp003621
18. MacLarnon AM, Hewitt GP. The
evolution of human speech: the role of enhanced breathing control. Am J Phys
Anthropol 1999;109:341-63.
https://doi.org/10.1002/(SICI)1096-8644(199907)109:3<341::AID-AJPA5>3.0.CO;2-2
19. Sundin L, Burleson ML,
Sanchez AP, et al. Respiratory chemoreceptor function in vertebrates
comparative and evolutionary aspects. Integr Comp Biol
2007;47:592-600. https://doi.org/10.1093/icb/icm076
20. Feldman JL, Del Negro CA,
Gray PA. Understanding the Rhythm of Breathing: So Near, Yet So
Far. Annl Rev Physiol 2013;75:423-52.
https://doi.org/10.1146/annurev-physiol-040510-130049
21. Hilaire G, Pásaro R.
Genesis and control of the respiratory rhythm in adult mammals. News Physiol
Sci 2003;18:23-8.
https://doi.org/10.1152/nips.01406.2002
22. Bianchi AL, Gestreau C. The
brainstem respiratory netÂwork: an overview of a half century of research.
Respir Physiol Neurobiol 2009;168:4-12.
https://doi.org/10.1016/j.resp.2009.04.019
23. Onimaru H, Homma I. A novel functional neuron group for respiratory rhythm generation
in the ventral medulla. J Neurosci
2003;23:1478-86. https://doi.org/10.1523/JNEUÂROSCI.23-04-01478.2003
24. Feldman JL, Del Negro CA. Looking for inspiration: new perspectives on
respiratory rhythm. Nat Rev Neurosci 2006;7:232-41. 10.1038/nrn1871.
https://doi.org/10.1038/nrn1871
25. Evans KC, Shea SA, Saykin AJ.
Functional MRI localization of central nervous system regions associated with
volitional inspiration in humans. J Physiol 1999; 520 Pt 2:383-92.
https://doi.org/10.1111/j.1469-7793.1999.00383.x
26. Nakayama T, Fujii Y, Suzuki
K. et al. The primary moÂtor area for voluntary diaphragmatic
motion identified by high field fMRI. J Neurol 2004;251:730-5.
https://doi.org/10.1007/s00415-004-0413-4.
27. Silverman WA, Andersen DH. A controlled clinical trial of effects of water mist on obstructive
respiratory signs, death rate and necropsy findings among premature infants.
Pediatrics 1956;17:1-10.
28. Mas A, Zorrilla JG, García D, et al. Utilidad
de la detecÂción del aleteo nasal en la valoración de la gravedad
de la disnea. Med Intens 2010;34:182-7. https://doi.org/10.1016/j.medin.2009.09.008
29. Giraldo BF, Chaparro JA,
Caminal P, et al. CharacterizaÂtion of the respiratory pattern variability of
patients with different pressure support levels. Conf Proc IEEE Eng Med Biol
Soc 2013;2013:3849-52. https://doi.org/10.1109/EMBC.2013.6610384
30. Yartsev
A. Alpha-stat and pH-stat models of blood gas interÂpretation, In Deranged
Physiology. A free online resource for Intensive Care
Medicine. Last updated Tue, 02/08/2022.
https://derangedphysiology.com/main/cicm-primary-exam/required-reading/acid-base-physiology/Chapter%20115/alpha-stat-and-ph-stat-models-blood-gas-interpretation
(accessed 18 June 2022).
31. Williams JJ, Marshall BE. A
Fresh Look at an Old Question. Anesthesiology 1982;56: 1-2.
https://doi.org/10.1097/00000542-198201000-00001
32. De Vito EL. Hipotermia [Hypothermia].
Medicina (B Aires). 1987;47:214-6. Spanish.
33. Karr A. Alphonse Karr : âPlus
ça change, plus câest la même choseâ. LâHistoire en citations,
https://www.histoire-en-citations.fr/citations/Karr-plus-ca-change-plus-c-est-la-meme-chose
2016 (accessed 18 June 2022).
34. Clark J. Temperature
Dependence of the pH of pure WaÂter. Chemestry, Libre Texts. Last updated Aug
15, 2020. https://batch.libretexts.org/print/url=https://chem.libreÂtexts.org/Bookshelves/Physical_and_Theoretical_Chemistry_Textbook_Maps/Supplemental_Modules_(Physical_and_Theoretical_Chemistry) /Acids_and_Bases/Acids_and_Bases_in_Aqueous_Solutions/The_pH_Scale/Temperature_Dependence_of_the_pH_of_pure_Water.pdf
(accessed 19 June 2022)

| GalerĂa de imĂĄgenes | ||
| Mujer joven con afectaciĂłn pulmonar bilateral y alteraciĂłn de la conciencia | ||
Autores: Churin Lisandro |
 |
|